|
MODULE II: CONTAMINANT BIOREMEDIATION
Session II-7
Bioremediation of Heavy Metals and Radionuclides
Previous Session | Table of Contents
Introduction. This session focuses on the ability of microorganisms to remediate heavy metal and radionuclide contamination. This remediation can occur using one or both of two different microbially-mediated mechanisms: (1) biosorption and (2) changes in the metal or radionuclide redox state via microbially induced oxidization or reduction. Biosorption is a physical phenomenon that results in concentration of the metal/radionuclide into the microbial biomass. Oxidation/reduction is biochemically based and results in a change in the metal/radionuclide's valence state that in turn can effect mobility of the metal/radionuclide in the subsurface. This change in valence state can be catalyzed directly by a number of different microbes as a part of their normal metabolic processes, or indirectly by interactions between the metal/radionuclide and microbially produced chemical products and/or intermediate electron carriers such as humics. Needless to say, unlike examples from previous sessions in this course, the microbes are not capable of degrading metals and radionuclides, but exert their effect by concentrating them or affecting their mobility through oxidation/reduction reactions.
Reading Assignment. Please read the following overview articles from the literature:
(1) Ledin, M. 2000. Accumulation of metals by microorganisms - processes and importance for soil systems. Earth-Science Reviews 51:1-31.
(2) Lloyd, J.R. and D. R. Lovley. 2001. Microbial detoxification of metals and radionuclides. Current Opinion in Biotechnology. 12:248-253.
(3) Lovley, D.R., and J. D. Coates. 1997. Bioremediation of metal contamination. Current Opinion in Biotechnology. 8:285-289.
(4) Lovley, D.R. and J. D. Coates. 2000. Novel forms of anaerobic respiration of environmental relevance. Current Opinion in Microbiology 3:252-256.
Pay attention to the particular metals and radionuclides that can be remediated using microorganisms, the species of microbes involved, and the chemical/physical conditions under which the remediation occurs.
Biosorption. Biosorption has been studied extensively over the last several decades. It is a property of certain types of microbial biomass, and relates to the ability of that biomass to sorb and concentration, independent of metabolism, metals and radionuclides. Since biosorption is independent of metabolism, both living and dead microbes can exhibit the phenomenon, and in fact in many instances, dead biomass may sorb more metals/radionuclide than its living counterpart. Biosorption includes both adsorption (accumulation at a surface) and absorption (accumulation through a phase). Biosorption is generally associated with the microbial cell wall or extracellular polymers (i.e. capsules or slime layers) exuded by microbial cells. Biosorption is usually a rapid phenomenon and generally can be described mathematically for single solute systems using Freundlich, Langmuir, or Brunauer- Emmett-Teller isotherms. All three of these approaches have different strengthens and weaknesses, and caution needs to be exercised when using the results from these isotherms, as they can be descriptive for a given system under a given set of conditions, but the results often can not be extrapolated to other systems.
Numerous metals and radionuclides can be biosorbed (Table 1) to high concentrations. As a result, it has been proposed that stimulation of indigenous bacteria so that they grow and produce copious quantities of extracellular polymers may be a strategy for immobilizing heavy metals and radionuclides in the subsurface. This could be accomplished by either immobilizing metals/radionuclides in the vadose zone to prevent contamination of underlying water supplies, or forming permeable biobarriers in aquifiers to remove metal/radionuclide contaminants from groundwater as it flows through the biobarrier.
|
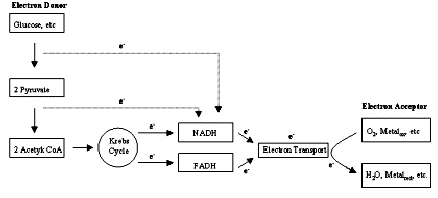
Direct Metabolic Reduction of Metals and Radionuclides. Microorganisms found in the subsurface are predominantly chemotrophs. Chemotrophs derive their cellular energy through the oxidation of chemicals, either organic or inorganic, as opposed to phototrophs that derive cellular energy from light. The chemicals that are oxidized for growth by chemotrophs are often termed electron donors, and as implied by this term, oxidation of a chemical by a chemotroph involves the removal of electrons from that chemical. For a microbe's metabolism to function, this oxidation ultimately must be accompanied by the concurrent reduction of an electron acceptor. Many compounds can function as electron acceptors including oxygen, nitrate, sulfate, carbon dioxide, and oxidized organic molecules like pyruvate. In addition, some microorganisms are able to use oxidized forms of metals and/or radionuclides as electron acceptors.
In some instances, the growth of a microorganism can be supported using these metals/radionuclides as a sole electron acceptor. In other cases, while the metals/radionuclides can accept electrons from a microbe and be reduced, growth only occurs if a second, non-metal/radionuclide electron acceptor is present. In either instance, the capability of microbes to metabolically transfer electrons to a metal/radionculide forms the basis for the direct metabolic reduction of metals and radionuclides as shown schematically in Figure 1.
|
Research over the past decade has demonstrated that a number of metals and radionuclides can be reduced by microorganisms. These include those shown in Table 2.
Iron is among the most prevalent of metals found in many subsurface environments. A variety of microorganisms including extremophiles (i.e. microbes that live at extremes of temperature, pH, etc,) can reduce Fe(III) to Fe(II), primarily under anaerobic conditions (Table 3). These organisms play a role in natural iron cycling, associated mineral formation, and decomposition of organic matter in aquatic, marine, and subsurface environments. They are also important for bioremediation purposes since many can reduce not only iron, but various heavy metals and radionuclides that are of environmental significance including Cr(VI), U(VI), Tc(VII), and Se(VI/IV/0).
|
|
A number of electron donors will support Fe(III) reducing microbial populations. Among these are acetate, lactate, formate, pyruvate, hydrogen, elemental sulfur, various fatty acids and alcohols. Certain organic environmental contaminants like benzene, toluene, phenol, p-cresol can also serve as electron donors for microbial Fe(III) reduction. As such, Fe(III) reducing microbes potentially can degrade these organic contaminants via oxidation while simultaneously reducing metals/radionuclides.
From a biochemical perspective, Fe(III) reducing microbes appear to use a number of different physiological mechanisms to transfer electrons to Fe(III). Among these are membrane bound iron reductases and c-type cytochromes.
Not all Fe(III) reducing microbes are able to generate usable cellular energy via Fe(III) reduction. However, some bacteria, under certain conditions can generate substantial energy using Fe(III) reduction. For example, when acetate is oxidized the standard free energy change at pH 7 is:
-193.4
kcal for Fe(III)
-201.0 kcal
for oxygen
-5.5 kcal for
Fe(OH)3
As such, Fe(III) offers energetics very similar to those obtained if oxygen is used as an electron acceptor. Conversely, this information illustrates that undissolved iron oxides and hydroxides (i.e. Fe(OH)3) are unfavorable electron acceptors at neutral or alkaline pHs. This has practical implications for the bioremediation specialist in that it might be difficult, if not impossible to establish a substantial Fe(III) reducing microbial population in the subsurface under these conditions without supplying a suitable alternative electron acceptor that the microbes can use to generate the bulk of their cellular energy. The choice of this alternate electron acceptor may vary from location to location and the most expedient way to select one is through laboratory testing using cores with indigenous microbial populations taken from the area of interest.
Cr(VI) is a toxic metal that is highly water soluble and mobile in the subsurface environment, however, its reduction product, Cr(III), is considerably less toxic, less water soluble, and less mobile. Microbial Cr (VI) reduction has been known for over two decades, with early studies showing that facultatively anaerobic Pseudomonas spp. are capable of catalyzing direct metabolic reductions of Cr (VI) to Cr (III). Since that time, numerous investigators have shown that the ability of bacteria to reduce Cr (VI) is a widespread trait across a number of chemotrophic and phototrophic bacterial genera as noted in Table 6.
Direct microbial reduction of Cr (VI) can occur either anaerobically or aerobically depending on the particular species or consortium in question. In addition, some species are capable of reducing Cr (VI) both aerobically and anaerobically depending on the redox potential of their growth environment.
While Cr (VI) reducing bacteria have been found in many Cr (VI) contaminated environments, the ability to reduce Cr (VI) appears to be independent of prior exposure to Cr (VI), with bacterial isolates from pristine environments exhibiting an ability to reduce Cr (VI) comparable to that observed with isolates from Cr (VI) contaminated areas. Furthermore, like the Fe(III) reducing microbes, the physiological mechanisms involved in Cr (VI) reduction appear to vary widely among species. In some cases, the enzymes catalyzing the reaction are membrane associated, while in other cases, the enzymes are in the soluble cell fraction. Many Cr (VI) reducing bacteria precipitate Cr extracellularly, others like Pseudomonas stutzeri have shown intracellular accumulations of Cr. As would be anticipated by the metabolically diverse genera listed in Table 4, a number of electron donors can be supplied to Cr(VI)-reducing bacteria with molasses, based on cost and availability being one of the best.
|
In a similar manner, a number of different microbes are capable of reducing uranium. Those shown in Table 5 reduce soluble, mobile U(VI) to less soluble, less mobile U(IV). Like Cr(VI) reduction, U(VI) reduction can be catalyzed using a number of different electron donors. These include short-chained organic acids and hydrogen. Many physiologically diverse reduction mechanisms seem to exist for U(VI) reduction. One of the best studied is reduction by Desulfovibrio vulgaris using ctyochrome c3.
|
Radionuclides other than U(VI) can be directly reduced by microorganisms. An example is Tc(VII) which can be microbially reduced to Tc(V) or Tc(IV). Up until now, the examples of metal and radionuclide reduction that we have considered have resulted in a soluble, mobile, oxidized metal/radionculide being reduced to a less soluble, less mobile valences. In the case of Tc(VII) reduction the opposite is true. Tc(VII) is less soluble and less mobile than either Tc(V) or Tc(IV), its reduction products, so in the subsurface, microbial reduction could potentially mobilize a relatively immobile Tc(VII) plume. Tc(VII) reducing microbes are shown in Table 6. As you examine the species listed in Table 6 you will notice that many that have been previously identified in this session as being capable of reducing metals or radionuclides other than Tc(VII).
|
In addition to potentially increasing solubility, microbially catalyzed metal reduction can also increase metal volatility. For example, a variety of microbes (Table 7) are capable of reducing Hg(II) to Hg(0) a significantly more volatile form of mercury. A least one enzyme system appears to be reduced in this reduction, a soluble NADPH-dependent, cytocplasmic flavoprotein. The activity of this system is not Hg(II) specific, but under laboratory conditions, has been shown to reduce ionic silver and ionic gold.
|
While up until now, we have been considering the impact of microbes on specific individual metals and radionuclides, the examples of Tc(VII) and Hg(II) reduction point to the need to consider the entire system approach when developing a plan to bioremediate a subsurface area. In many instances, metals or radionuclides are present with a mixture of other contaminants, both organic and inorganic. In effectively remediating an area, it is important to understand all contaminants present and to ascertain the effect of a given bioremediation strategy on the total system. It is possible to remediate one specific contaminant, but through that remediation have a net deleterious impact on the system by causing increases in the toxicity and mobility of non-target contaminants that may also be present.
Indirect Microbial Metal Reduction. Thus far, we have considered microbially induced metal reduction in the context of a direct transfer of electrons to a metal/radionculide via enzymatically catalyzed reactions that are part of the metal/radionuclide reducing microbes' normal physiology. Microbes are also capable of reducing metals indirectly either by the reaction of a metabolic byproduct or by transferring electrons via chemical intermediates.
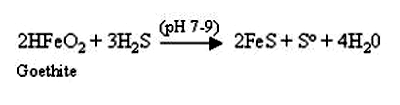
Perhaps the best known of the indirect metal reducing microbes are the sulfate reducing bacteria (SRB). While some SRB can directly reduce metals via the direct, enzyme mediated metabolic processes as discussed earlier in this session, all SRB share a common property of being able to grow anaerobically using sulfate as a terminal electron acceptor with sulfide, often in the form of hydrogen sulfide (H2S) being the end product of this reduction. Depending on the species of SRB in question organic acids, alcohols or hydrogen, which are often fermentation products from other microorganisms, can serve as electron donors. The hydrogen sulfide produced can react chemically with a variety of metals in soil. An example of this is shown in Figure 2 in which hydrogen sulfide is reacting with and reducing iron in the mineral goethite. Similar reactions can occur with other heavy metals such as chromium, lead, and cadmium, leading to their precipitation and immobilization.
|
The ability of metal reducing microbes to reduce metals via chemical intermediates has also been studied in some detail. The best known example is the ability of Fe(III) reducing bacteria, under anaerobic conditions, to use humics or humic analogs such as anthraquinone -2,6-disulfonate (AQDS) as electron acceptors. Once microbially reduced, these humics can then abiotically transfer electrons to metals such as Fe(III) which is in turn reduced to Fe(II). Studies suggest that the electron accepting moieties in humics are probably quinones, and there appears to be a direct correlation between quinone content in humics and their electron accepting capabilities. It has been recently demonstrated that the addition of humics or humic analogs to Fe(III) reducing bacterial cultures can facilitate the reduction of iron minerals that the bacteria can not reduce directly. As such, evaluation of potential subsurface bioremediation sites should consider the presence of humics since they may enhance microbially catalyzed metal reduction.
Oxidation of Metals and Radionuclides. Many microorganisms can catalyze the oxidation of metals such as iron, manganese, selenium, tellurium, and various metal sulfides. Some of these microorganisms, termed chemolithotrophics gain their cellular energy through the oxidation metals. Among these, iron oxidation has been the best studied, with a variety of bacterial indigenous to soil and the subsurface having been shown to oxidize ferrous iron (Fe(II)) to ferric iron (Fe(III)) which results in a gain in cellular energy for these bacteria. Included among these iron-oxidizing genera of bacteria are Thiobacillus, Leptospirillum, and Metallogenium. While some of these organisms can grow under both aerobic and anaerobic conditions, they typically tend to use oxygen as a preferential electron acceptor when oxidizing metals. As discussed above, the changes in valence state catalyzed by these bacteria can effect metal mobility and toxicity.
Many of these same organisms can affect metal mobilization by bioleaching. Bioleaching depends on the biological formation of acids that then leach metals from soils. This process has been used (albeit not understood) since Roman times to mobilize and concentrate copper from low grade ores. Iron and sulfur oxidizing chemolithotrophic bacteria like Thiobacillus ferrooxidans and Thiobacillus thiooxidans can be enriched from contaminated soils and used to leach through biogenic formation of sulfuric acid, metal contaminants from those soils. Common metal contaminants subject to leaching include Cu, Cd, Co, Ni, Zn, V, Pb, and As.
Acidogenic chemoorganotrophs (bacteria that form acids while chemically oxidizing organic compounds) also may be useful in leaching metals and radionuclides from soils. These organisms produce organic acids and are capable of growing a neutral and alkaline pH's as opposed to the chemolithotrophs that typically grow at acid pH's. As such, the use of chemoorganotrophic leaching may be appropriate for many more contaminated soils with pH's above 5.5-6.0 than is chemolithotrophic leaching.
Questions
Describe the difference between chemoorganotrophic, chemolithrotrophic, and phototrophic microbes. Which would be more useful in the bioremediation of subsurface soils?
Describe three different bioremediation methods for the immobilization of metals/radionculides.
What are the differences in metabolic energy requirements for biosorption, bioreduction and bioleaching? Do all require an exogenous energy supply to function?
Describe the effects of reduction and oxidation on metal mobility. Give examples of metals/radionuclides that are immobilized when reduced. Give examples of metals/radionuclides that are mobilized upon reduction.
Homework
The Hocking Valley is a 2 mile long valley with an average width of 1/4 mile. The elevation change between the upper and lower end of the valley is approximately 750 feet. Sapphire Creek originates from springs and snow run off at the upper of the valley, runs the length of the valley where it empties into the Bristol River. The valley originally was forested, primarily with stands of aspens and Douglas firs, most of which had been cut in the last 3 years. Several large piles of sawdust are spaced around the valley as a result of these logging operations. From the mid-1800's to the early 1900's a series of mining operations were conducted in the valley, initially for silver, and later for copper. These mining operations were primarily small, family-owned businesses and consisted of drift mines dug into the faces of the valley's steep hillsides. There are 23 of these drift mines in the valley, several of which were dug to depths of 500-750 feet as the miners followed individual veins of metal-bearing ores. Metal-bearing groundwater now flows from the openings of these drift mines, down the hillsides and into Sapphire Creek. A recent analysis of water from Sapphire Creek has shown the pH to be 2.3, and to contain 1.4 mg/L Pb, and 180 mg/L Cu, 1200 mg/L Fe, 2.2 mg/L Cd, and 2200 mg/L SO42-. Sapphire Creek water oxygen levels average 4 ppm(v) and the streambed is coated with a yellow precipitant consisting primarily of ferric hydroxy sulfate complexes. Beyond microorganisms there is no life in Sapphire Creek and drainage from Sapphire Creek has contaminated the Bristol River, adversely affecting its sport trout fishery for a distance of 20 miles downstream from the confluence.
You have been hired to remediate Sapphire Creek and save the trout fishery in the Bristol River. Design a bioremediation program to do so. Keep in mind both efficacy and cost considerations as you design this system.