Biofilm Growth and Development
From the microbial point of view, being included in a biofilm is a very good thing. Here we will discuss the development of typical biofilms and the advantages of being included in such a structure. It is now clear that biofilms possess properties that could not be predicted from a detailed understanding of the individual microorganisms of which they are constructed. These emergent properties, like, complex architecture and resistance to antimicrobial agents, for example, are the key attributes that make biofilms both interesting to investigate and essential topics for research due to their beneficial and inimical effects on human culture. In this chapter we will explore the developmental biology of biofilms, the advantages to a bacterium of being part of a biofilm and, those emergent properties which make biofilms unique.
The Developmental Biology of Biofilms
The Conditioning Film
Within moments of placing a clean sterile slide into a water source, be it a pond, stream or an aquarium, a film begins to form that consists of proteins and polysaccharide molecules adsorbed to the glass surface. Similar films develop on a professionally cleaned tooth surfaces or on the surface of indwelling urinary catheters. It is to this so-called ‘conditioning film’ that the first cells of the insipient biofilm attach. Such conditioning films begin to form so rapidly, perhaps in seconds, that there is a question as to whether bacteria can attach to a barren substrate lacking such a film. In any case, the accumulation of the conditioning film continues to develop for hours following first exposure.

Note that when placed into sterile tissue, the conditioning film develops even though bacteria are absent.
Conditioning films form on both hydrophilic (e.g. glass) and hydrophobic (e.g. many plastics) surfaces. A great deal of effort time and money has been invested by industries attempting to find surfaces to which biofilms will not adhere. Such a material would be of enormous value in many industrial applications including water and oil pipeline manufacturing, heat exchangers, air conditioning systems and also in medical applications such as indwelling catheters, cardiac pacemaker probes, prosthetic implants (hip, knee joints) and hydraulic dental drills. To date, no surface has been discovered that will reliably inhibit the formation of biofilms, although mistakes, costly in lawyer's fees and human lives, have been made on the assumption that such materials had been found. Instant fame and fortune await the individual or corporation that solves the mystery of the biofilm free surface material.
Bacterial Approach and Contact
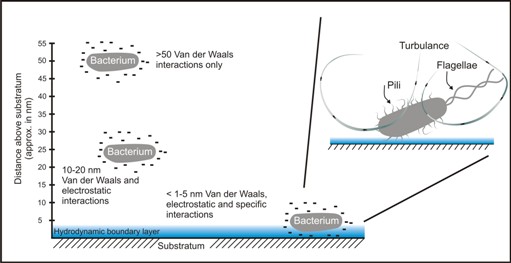
Two forces act on a bacterium suspended in a fluid stream close to a substratum wall (Figure 2). The first, the Van der Waals force, is attractive and acts at a distance of a few hundred nm (a few tenths of a mm). This force tends to bring small objects close to the substrate wall. The second force is due to the fact that bacterial cells and many environmental surfaces possess a net negative electrostatic charge. These negative charges produce a repulsive force as the bacterium approaches the surface at a distance of about 10-20 nm. When the bacterial cell comes to within about 5 nm the probability of physical contact by cell surface receptors, such as pili and flagellae and their associated adhesins, increases greatly.
It is also in this layer close to the substratum wall that flow velocity is much reduced. This region called the hydrodynamic boundary layer is caused by drag and frictional forces and provides a low shear, low turbulence area in which physical contact between the bacterium and the wall may occur. Physical contact with the conditioning film may bring about a transient and potentially reversible attachment to the substrate.
Any increase in kinetic energy may boost the probability of a bacterium overcoming the electrostatic force and this may be a major function of bacterial flagellae. In addition, any projections such as pili that extend through the zone of electrostatic repulsion may increase the likelihood of bacterial attachment to the substratum wall (Figure 3).
Reversible Attachment
Video photo-micrography has been of enormous importance in deciphering the events surrounding the attachment of bacteria to surfaces. The best evidence suggests that attachment occurs as a two-step process. Reversible attachment, which is tenuous and often transient, and irreversible attachment that is much more stable.
In some cases, following their initial attachment to a surface the bacteria can be seen to be rapidly spinning, vibrating or actually moving across the substrate surface. Spinning is an indication that the cells are attached to the substrate by their flagella, but since the flagellum is now “fixed”, the cell body rotates as the flagellar motor continues to rotate. Sometimes, cells in contact with the surface seem to be vibrating, that is, they are exhibiting Brownian motion, caused by the constant and random impact of water molecules striking the loosely attached bacterial cell.
Click image shown above to view video
In this film (Video 1) of Xylella fastidiosa, bacteria can be seen attaching to the substratum surface even in a flowing environment. Some of these subsequently detach and “go with the flow” down stream (Left to right). Other cells in contact with the surface exhibit “twitching motion” which enables them to move from right to left against the fluid flow (red circles).
The cells of some organisms, once they have reversibly attached to a surface can move about by means of what is called “twitching motility” (Video 2). In this novel mode of locomotion, the cells extend a pilus which, attaches at its tip to the substratum surface. The pilus is then contracted, pulling the cell forward in a twitching jerky motion. The process is then repeated.

Click image shown above to view video
As can be seen in Video 3, the movement can be quite rapid. The cells shown here are Xylella fastidiosa moving at the margin of a larger colony.
Click image shown above to view video
Twitching motility also plays a role in the formation of microcolonies as the biofilm begins to take shape. Individual cells autoaggregate to form colonies with what will remain as water channels between (Video 4). In this video the cells actively aggregate. At intervals a large mass of bacterial cells which have broken away from the surface disrupt the aggregation process which begins anew.

Click image shown above to view video
Irreversible Attachment
At some point following initial contact, the cells become encased in a polymeric matrix that the cells produce and thus are anchored to the substratum. Although the time required to transition from reversible to irreversible varies greatly depending on the organism being observed and the physical conditions present it is generally on the order of minutes, not seconds or hours. The more stable attachment is due to the production by the bacteria of a complex array of extracellular polymeric substances including polysaccharides, proteins and in some cases, DNA.
The use of reporter genes such as the Green Fluorescent Protein placed in the bacterial genome just behind the promoter region of the native gene or operon indicate that attachment causes the quorum sensing induction of genes responsible for the production of compounds found in the matrix of developing biofilms such as the genes of the alginate pathway. Alginate, a polysaccharide, is a major component of matrix EPS (Extracellular Polymeric Substance) in Pseudomonas aeruginosa, an important opportunistic pathogen. The genes responsible for the synthesis of alginate are turned on only after the bacterium has formed an irreversible attachment to the substratum. In fact it has been shown that attachment causes a major change in phenotypic expression in biofilm forming organisms that may involve a change in expression of as much as 70% of the genome.
